��������
�ذ�����حά����D��л�방֢��ϵ
ǰ��
ά����D����ͳ�ϱ���Ϊ��һ�ֱ����֬����ά���أ����Դ���ʳ��ã�Ҳ���Ծ��������������Ƥ���кϳɣ��ǹ̴����غϳɵ�ǰ�壬�㷺����������������̡�ά����D�ľ����л·�����ڸ���������о���������л������һ�־����������ԵĴ�л��ǻ����������ܹ���ά����D���壨VDR����ʹ�����������ܣ�����Ľ�ɫ�ǵ��ڹ��ؽ��еĸƺ��״�л��Ȼ�������ڵ��о�������Ҳ���Ա�һ��CYP11A1��ʹ�ķǵ��ʹ�л·����л�ͼ��
Ƥ��ȱ����������ʳ���䲻����������ά����Dȱ���������о���������ȱ�������ӳ��˹���������������������簩֢�ķ������ա��״ι۲쵽���ղ���Ͱ�֢�ķ����������ʳ�����ع�ϵ����80��ǰ�ı����������1980���1992�꣬��һ�������в�ѧ�о������չⱩ¶�ͽ�ֱ������ǰ���ٰ��ĸ߷����ʽ�������������ά����D��Ϊ���յ�һ���������Խ��ͽ�ֱ������ǰ���ٰ��ķ������ա������Ժܶ����в�ѧ���о���֧�����漰18�ְ�֢��UVB-ά����D-��֢��˵�����ּ�˵���ά����D�Ͱ�֢���յ�ֱ��������о�֤ʵ�����⣬�ܶ���Ⱥ�����о���ʾ��Ѫ���е�25-�ǻ�ά����D (25(OH)D)ˮƽ�ͽ�ֱ���������ٰ���ǰ���ٰ���θ����������һЩ��֢�߷��ճʸ���ء����ң�һЩϸ���Ͷ���ʵ��ҲΪά����D�Ŀ��������ṩ��ǿ������֤�ݡ���ˣ�ά����D��ȱ���ܴٽ��֢ܶ�ķ����ͽ�չ��ά���㹻��ά����Dˮƽ������Ԥ�������ư�֢��
��Ȼ������в�ѧ�о���ʵ�����ݱ�����ά����D��Ԥ�������ư�֢�е����ã���ô���Ƕ��ڹǻ�������ά�������������ٴ��е�Ӧ��Ҳ�������о���Ȼ��������ʹ��ά����D�����ĸ�Ѫ��֢�����������ٴ��е�ʹ�á����ң����㹻�����ݱ�����ϸ��ͨ�����ַ�ʽ���ǻ�������ˮƽ�������书�ܼ����Ϳ������ԵĽ��͡���ˣ��о�ά����D�ڰ�֢����δ�л����лʧ�������ڿ˷�Ŀǰά����D�������������ٴ���Ӧ���е����ơ�
ά����D��л
ά����D��Ϊһ�ּ���ǰ����Ҫ��л�ɾ����������ԵIJ����ͬԴ��������ȥʵ����Բ�ͬ�������ܵĵ��ء�
�������
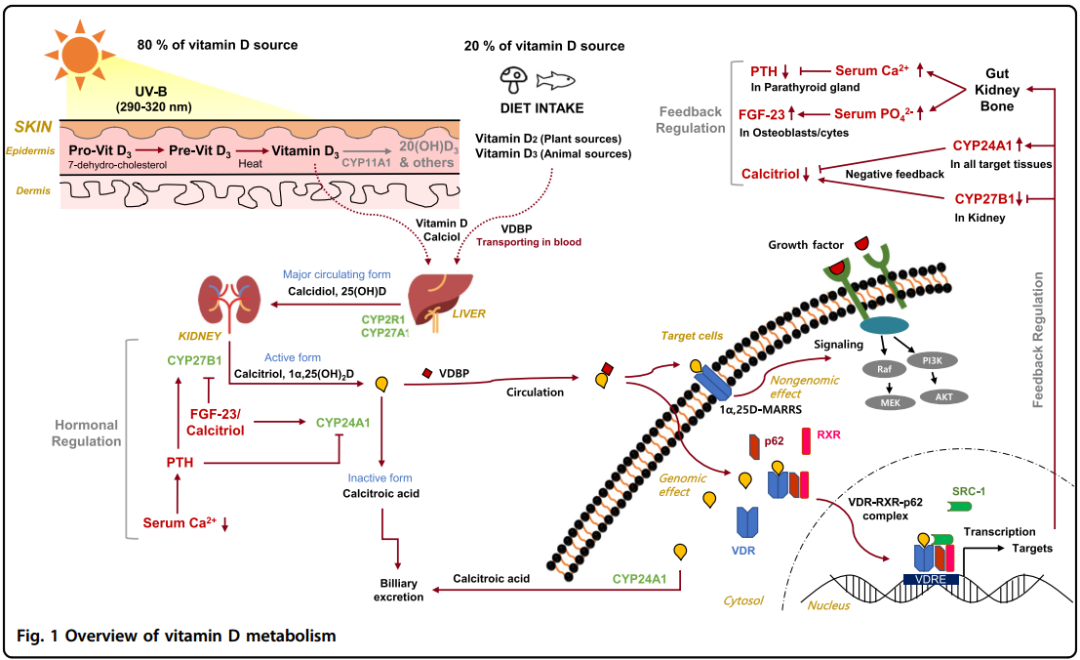
ͼ1ά����D��л����
ͼ1������ά����D�ľ����л·����ά����D��������Ҫ���칹�壬ά����D2����Ǹƻ�������ά����D3�����ƻ�������ά����D2��ֲ���ĸ������к��е���ǹ̴��������������ϳɣ�Ҳ���Դ�ֲ��Դ�Ե�ʳ������Ģ�������롣ά����D3�ɱ�Ƥ�е�7-����̴������������ϳɣ�Ҳ����ͨ������Դ�Ե�ʳ����������������롣�����Ǵ���ʳ����Ƥ���л�õ�ά����D2��ά����D3��Ҫ���Ȱ�ά����D-��ϵ��ף�VDBP������ѭ����Ȼ���ٱ����䵽���ࡣ
��������ά����D��25-�ǻ�ø��CYP2R1��CYP27A1����л��25(OH)D���ǻ���������25(OH)D��Ѫ����ά����D����Ҫѭ����ʽ��25(OH)D����������С���е�1��-�ǻ�ø��CYP27B1����л��1��,25(OH)2D���ǻ�����������ά����D������������Եijɷ֡��ǻ���������ѭ�����VDBP�����䵽�����ǡ������Ŀ����֯�У���������ڸƺ������ա�ת�ƺ������ա�һ�����ɣ��ǻ������ǻ�������ˮƽ�ͻᱻ25(OH)D
24-�ǻ�ø��CYP24A1���ϸ���أ�����ø���ǻ������ǻ�������C-24��C-23λ�ǻ�����24-�ǻ���·��������л������ᣬ������û�������������մӵ�֭�ų����⣬�����˽���ٵ�23-�ǻ���·�����γ�1,25-26,23��֬������·���е�CYP24A1�����Ա��ֳ����ֲ��졣CYP24A1�ǻ��������ջ�ʹά����Dʧȥ���������������һ���о��б����ֵġ�������о��У�Cyp24a1���ó���С����ֳ�Ĥ�ڹ������Ѫ��֢��������50%��С����������Ȥ���ǣ���Cyp24a1��VDR�����ó���С����û������ȱ�ݣ������ǻ����������Ӷ�����24��23-�ǻ���ά����D��л���ȱ������������ȱ�ݱ��͵IJ�����
��Ŀ����֯�У��ǻ�������VDR��VDR�����弤��ת¼���Ӻ���������е�һ�֣�������Ը��������������ܵĻ����ͺͷǻ����͵ĵ��ء��ڻ���ͨ·�У��ǻ�������ϸ����VDR��ʹVDR���ữ��Ȼ���RXR�γ���Դ�����壬���Ը�������к�ת�ˡ��ǻ������CVDR�CRXR�������Ŀ������������е�ά����D���嵥Ԫ(VDRE)����ļת¼�������ӻ�����������ȥ����Ŀ�����mRNA�ı�������ڲ�ͬ���������ܣ������ƺ��Ĵ�л�ȡ���Ȥ���ǣ�һ����ڵ��о������������䵰��p62/SQSTM1ͨ���ڸ���״ϸ����ֱ�Ӱ�VDR��RXR�ķ�ʽӰ��VDR-RXR�γ���Դ������Ͷ�Ŀ��������ļ���ڷǻ���ͨ·�У��ǻ�������Ĥ��ϵ�VDR������VDR��һ�ֿ�����Ӧ����̴�-����(1,25D-MARRS)������������������ϸ���ź�ͨ·�ľ��ұ仯�������ƺͷ�����-�����ø(MAPK)���ź�ͨ·��ͨ����ϸ�����źŷ��Ӽ��ֱ�ӵ���-�������ò����ijЩ�����ܵĵ��ء�
�������
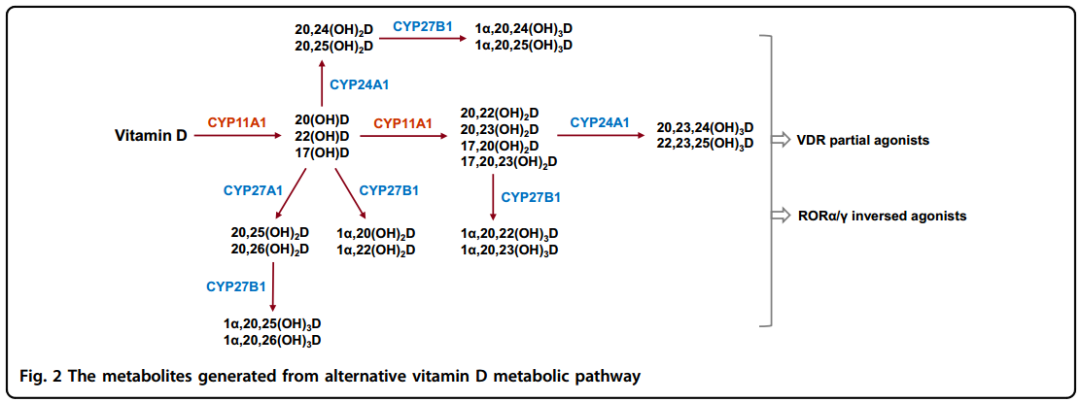
ͼ2ά����D�����л·��
��ͼ2����������ϸ��ɫ��P450�����ѽ�øCYP11A1�����µ�ά����D�������л·��������������CYP11A1���������̴��������д���̴����ɵĵ�һ������ø����ԭ���������յ�C-22��C-20λ�ĵ��̴��ǻ��������ʹC-20��C-22֮��ļ����Ѳ�����ϩ��ͪ����̴����صij���ǰ�壩����ô����ô����CYP11A1��һ���µ�ά����D��лø���أ��о�����CYP11A1����Ե��֯����Ƥ����θ�����б��ά����D����������̴���Ϊ����ø�ĵ��CYP11A1����ά����D��C-20��C-22λ���������ǻ��������Ҳ��ᷢ�������Ķ��ѡ�CYP11A1���ĵ��ǻ���������Ҫ��20(OH)D��22(OH)D��17(OH)D����Щ�ǻ����IJ������һ����CYP11A1�ǻ����γ�20,23(OH)2D��20,22(OH)2D��17,20(OH)2D��17,20,23(OH)3D�����ң�����·������Ҫ����20(OH)DҲ����ΪCYP27A1��CYP24A1�ĵ��CYP27A1�����µ��ǻ���λ����C-25��C-26,��CYP24A1�ǻ�����λ����C-24��C-25����CYP27B1�Ĵ�����Щ�����C-1��λ�û����Ա��ǻ����������ǻ���ά����D��л���Щ��л�ﲻ����17,20,23(OH)3D������CYP11A1��ʹ��ά����D��л�ղ����֮�����Ʋ���������Ĵ�л·�������ɳ���21��ά����D���ǻ�����л���
��CYP11A1���IJ����У���20(OH)D�������ǻ�������Ϊ����������Ƥ��ϸ��������ǻ������൱���߸��õ��յ�����ֳ���ֻ��Ϳ���Ч�������ң���Щ��л��������յ��µ�DNA���˺�����Ӧ�����ֳ���ǿ�ķ������ƣ���һЩ���ϸ��ϵ�и��õĿ������ԡ���Ȥ���ǣ�CYP11A1�IJ������20(OH)D��20,23(OH)2D����VDR�в��ֻ�ƫ��ĵֿ����á��ٸ����ӣ���Щ��л���ڼ���VDR��ҩ��Ũ��ˮƽ�ϲ����յ�Ѫ��ЧӦ����CYP24A1�ı�����������������ڹǻ��������ƱȽϳ�����
����VDR,��ά����A�������(ROR��)�ͦ�(ROR��)����-��������ת¼���Ӻ���������еij�Ա��Ҳ������Ϊ20(OH)
D��20,23(OH)2D�µ����塣RORs������ںܶಡ֢���簩֢���������߲��ʹ�л�쳣�е����ߺʹ�л���̡���Ȥ���ǣ�CYP11A1������ά����D��л�������ΪROR����ROR�õķ�����������ת¼���ԡ��ٸ����ӣ�20(OH)D��20,23(OH)2D�ܹ��ֱ�����ROR����ROR��Ŀ�����Bmal1��G6Pase��ת¼����ˣ�ά����D�Ķ�Ч�Ժ���ЧӦ���������ڹǻ�����-VDR·����Ҳȡ����CYP11A1������ά����D��л��-VDR��-ROR��/��·������һ������Ҫ����Ĺ���������ά����D�Խ����ͼ����Ĺ㷺Ӱ�졣
����ά����D��л�ļ��ص���
�ǻ���������Ϊһ�ּ��أ���һ�ָ��������ƽ��ܵ���ά����D��л�У�ʹά����Dʧ���ø-CYP24A1���ǹǻ�����-VDR-RXR��������ǿ��ת¼Ŀ��֮һ��CYP24A1�������Ӱ���2���ӽ�ת¼��ʼ������150��250-bp��VDREs,�����ǻ�������CYP24A1��ǿ���յ������⣬ͨ����ļ�鵰��H4����ת��ø��λ������CYP24A1��������50��70-kb��RNA�ۺ�øII���ǻ�����Ҳ���յ�CYP24A1�ı����ˣ������йǻ�����������CYP24A1�����ܽ��ܵ��ڹǻ������ǻ�������ˮƽ�����ң��ǻ�����Ҳͨ��������������ı����Ŵ����εȸ��ӵĻ�������������CYP27B1��ת¼��
���˹ǻ������ĸ��������ڣ���״���ټ�����PTH���ͳ���άϸ���������ӣ�FGF-23�����ּ���Ҳ�ܵ���ά����D��л�������ּ��ض�ά�ָƺ�����̬������Ҫ���á�PTH�ɼ�״���ٷ��ڶԵ�Ѫ���Ũ������Ӧ���Ƹ��������壨CaSRs���ڼ�״����ϸ�����汻���PTHͨ��cAMP-������ת¼���ӻ����ϵ�����������NR4A2-������ת¼�̼��������CYP27B1����ʹ���ӹǻ��������յ�CYP24A1�ı����������������⣬PTHҲ�ܼ��������е�cAMP�CPKA·���յ�mRNA��CYP24A1�Ľ���ʹ�ǻ�������ˮƽ�õ�ά�֡�����ά�ֵ��µĸ�Ѫ��ˮƽ����һ�ָ��������ڻ���ͨ����״�����е�CaSRsʵ�ֶ�PTH���ڵĸ�����������FGF-23�ɳɹ�ϸ����ϸ�����ڣ��Ը�Ѫ�߸�ˮƽ����Ӧ��FGF-23��������С�ܽ��˶���Ĥ�ϵ���-��Эͬת�˵���2(NPT2)�ı���ٽ��ķ��ڣ���������ͨ����ϸ��Ĥ�е�FGF receptor�CKlotho������ʵ�֡����⣬FGF-23����������CYP27B1�ı���ͬʱ�̼�CYP24A1�ı���ʹѪ���еĹǻ�����ˮƽ���ͣ�Ȼ�����ֻ���Ŀǰ���д�֤ʵ��
ά����D�Ŀ�������
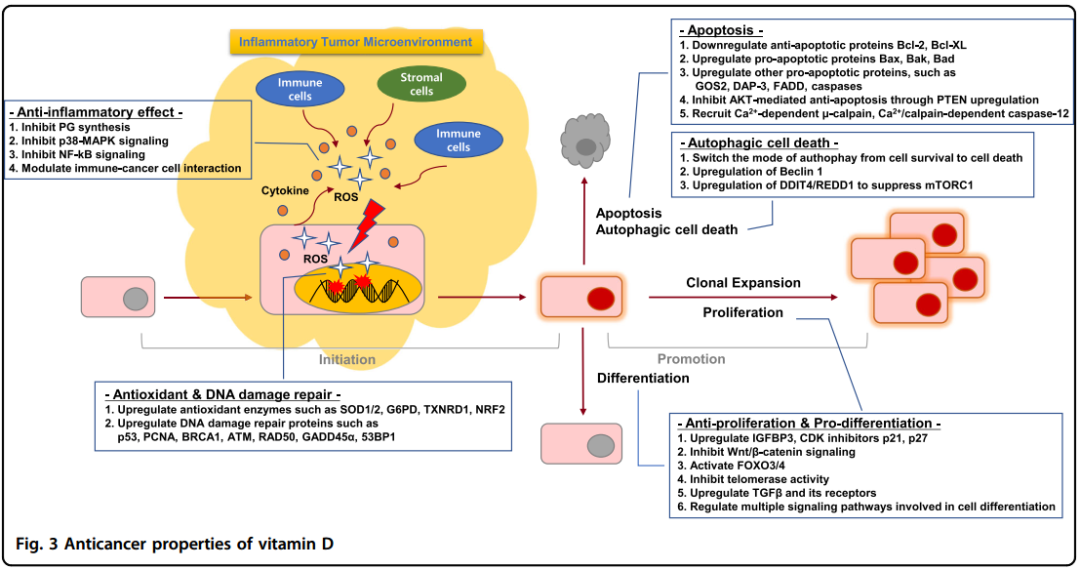
�����Ѿ������в�ѧ���ٴ�ǰ�о��۲쵽��ά����D��Ԥ�������ư�֢����Ļ���Ч��������Ҳ����˺ܶ������������һ�����д����о�������ʾά����D�ܹ����������������������̣����ʼ������ת���Լ�ϸ��-����������ã���ͼ3��ʾ����Щ������������ϸ������Ϊ��������ֳ���ֻ������������ɺ���Ƥ������(EMT)��ת�䣬�Լ�����ϸ��-��������ã�����Ѫ�����ɡ�����������֢������ϵͳ��
 ��
��
ͼ3ά����D�Ŀ�������
��ʼ�Σ��ڿ��ס���������DNA�������е�����
�����ij�ʼ����һ��������ϸ���в�����Ļ���ͻ����̣�����յ��˱仯���ܶ����ݱ���ά����D�������γɽ��ؼ����谭���ã���������ͨ�����ס��������Լ�DNA���������̵��Է��ӡ�
����
������֢��һ�ֵͳ̶ȵij�����Ӧ�����յ�����֯���������ڻ�������ROS������֢λ����ڵ�ϸ�����ӵ������½��������������ǹ㷺��ʶ��������֢�ǵ�������������һ����Ҫ���ء���ά����Dͨ������4�����Ʒ��ӿ������á�
��һ�ֻ����ǹǻ�����ͨ�����ƻ�����ø-2(COX-2)��PG����ı����Լ��ٽ�PGs�Ľ������ƴ�����Ӧ��ǰ������·������ǰ���ٰ�ϸ���У��ǻ�����������COX-2��PG����EP2��FP�ı��ͬʱ������15-�ǻ�ǰ��������ø(15-PGDH-һ��NAD+-�����ĸ���PGE2�����ø)�ı�����⣬�����ٰ����о��б������ǻ��������ƺ�COX-2mRNA�������ٺ�PGE2�����ӵġ������ٰ�ϸ��ϵ���ѳ�����֯��VDR�ı�����COX-2�ʸ���أ�֧���˹ǻ�����-VDR��������COX-2�����PGs������һ�۵㡣
�ڶ��ֻ�����ά����D��������p38
MAPK���ڵĴ���֢�ź�ͨ·����������ǰ������Ƥϸ����ǰ���ٰ�ϸ���У��ǻ������յ�MAPK����ø-5�ı������ƴ���֢ϸ����������IL-6�����ɣ���һ������ֹ��p38
MAPK�����ữ�ͼ������ǰ����ϸ�����ǻ�����Ҳ���յ����嵥��ϸ������ľ���ϸ���е�MKP-1������֬�����γ�IL-6��������������(TNF)-����
�����ֻ����ǹǻ������ܹ����ƺ�����(NF��B)�ź�ͨ·���ǻ��������ƾ���ϸ���е�AKT��������Ŀ����1
I��B�������ữ����������ͨ���ϵ�THEM4(һ������NF��B��AKT���ص���)��COX-2�ı������ʵ�֡��ڳ���άϸ���У��ǻ�����������I��B�������ȶ��ԣ�ͬʱ�յ�VDR��I��B��ø�İ���ֹ�����ữ�ͼ����������NF��B�ǵ�Ԫp65�ĺ�ת�ơ�
�����ֻ�����ά����D���Ե������ߺͰ�ϸ��֮�������ý������ƴ���ϸ�����ӵ����ɡ�����Ѫ����ϸ���ͽ�ֱ����ϸ��������ʵ�����ά����D���ƿ����������ʹ���ϸ����������TNF-��,
IL-6�͵ͳ̶ȵ�IL-10�����ɣ�֧����ά����D�����������еĿ���������һ�۵㡣
��������DNA������
ROS�����������ĺܶ�涼������Ҫ���ã����ܴٽ�DNAͻ�䡢ϸ����ֳ��ϸ����������ˣ�ά��ϵͳ�Ŀ��������Ƿ�ֹ������չ�Ĺؼ����衣Խ��Խ������ݱ���ά����D�ܹ��ٽ���������������Ӧ�����µ�DNA���ˡ��о�������VDR�ó���С��᳦��Ƥϸ������������Ӧ��������DNA�������������ơ����ң��ùǻ��������Ƶ�С�����������ͱ���ȩˮƽ������ȩͨ����֬�ʹ���������DNA���˵��ղ���ճ�����ά����D���Լ��ٹ��������µ�DNA���ˣ������ά����D�ܹ����ƹ�����Ӧ�����µ�DNA���ˡ�
ά����D���ڵĶ�ROS�յ���DNA���˵ı�����Ҫ��������������ROS�ⶾ�����д���ø�ı���о����֣��ǻ������ֱ��յ�ǰ������Ƥϸ�����������绯ø1(SOD1)���ۼ������е�ǰ���ٰ�ϸ���г������绯ø2(SOD2)�ı�����⣬�ǻ��������յ�������ԭ����ø1(TXNRD1)�ı������ø�ή��ǰ���ٺ����ٰ�ϸ����������ԭ���Ŀ��������ܣ��ǻ����������յ�������-6-��������ø�ı������ø��ǰ���ٺ��ѳ���ϸ���в���NADPH���ڹ����ĵ����������ң�NF-E2-�������2��NRF2��,һ���������Ӷ��ֿ�������ø�ı����ת¼���ӣ�ά����D����ͨ������NRF2�ı����ת�˻�����KEAP1���صĽ�����������е��ء�����NRF2�ǿ�����ø����Ҫ�����ߣ������Ҳ��ά����D�ܹ��յ�������ø�ͱ��ֿ��������õ�DZ�ڻ��ơ�
����ͨ���Ŵ���������Ԥ��DNA�����⣬ά����DҲ��ֱ�ӵ���DNA�����������̡��о�����ά����D���Ӳ���DNA�����������еĻ���������p35����ֳϸ���˿�ԭ(PCNA)�����ٰ�ϸ���е����ٻ���1(BRCA1)��PECs�е�ëϸ������ͻ�����(ATM)����ļDNA������(RAD50)����״��ϸ���е�����ͣ�ͺ�DNA�����յ���(GADD45��)��ά����DҲ����ֹ�����ٰ�ϸ���е���ø����������ø���װ�����֯����1ø���ڵ�p53-�ĵ���1(53BP1)�Ľ��⡣��ˣ�ά����Dһ���̶��Ͽ���ͨ���յ����ס���������DNA���������ܷ�ֹ������ʼ�εĻ���ͻ�䡣
��չ�Σ���ϸ����ֳ/�ֻ��͵���/�����е�����
��ʹϸ���Ѿ��ڳ�ʼ�η����˻���ͻ�䣬ά����D��Ȼ��ͨ������ϸ����ֳ���յ��ֻ����������谭������չ�������ӿ������á�
ϸ����ֳ�ͷֻ�
�ǻ�������֤���������Ͱ�ϸ���о�����ֹϸ����ֳ�ʹٽ��ֻ������á����ҳ��˹ǻ���������CYP11A1���������µ�ά����D��л������20(OH)D3��20(OH)D2��1,20(OH)2D3��20,23(OH)2D3����ͨ��VDR����ϸ����ֳ���յ��ֻ������ã�����Ч����ǻ������൱�����ǶԸƵ�Ӱ���С��ά����D�Ŀ���ֳЧ���ܵ����������������ӡ�ϸ��ѭ�����ź�ͨ·���ڵĶ��ֻ��Ƶĵ��ء���һ��ά����D���������ȵ����������Ӱ���3��IGF�������ڵ���������ø��CDK�����Ƽ�p21��p27�ı��ͬʱ����CDK2�ı����������IGF-1��IGF-2�̼���ϸ����ֳ��ϸ�����ڡ��ڶ����ǻ���������ת¼����4-��-���������γɻ�����Wnt����DKK-1�ı���������Wnt/��-���������ź�ͨ·��������ά����DҲ�ܼ���ת¼����FoxO3/4�����ܹ��̼�ϸ������ͣ�ͺͿ���ֳ��Ŀ������ת¼��ͨ��ʲô��ʽ�أ�ͨ���յ���ĸϸ����ϸ����ȥ��������ȥ���ữ��ɡ����ģ�ά����D�������ƶ���ø�Ļ��ԣ������յ�ת���������ӣ�TGF�£���������ı����������ϸ�����������壬ά����D�ܵ���ϸ���ֻ����漰�Ķ���ź�ͨ·����������������3��ø/AKT��MAPK��NF-kB��Ca2+�ź�ͨ·��
����������
ά����D�յ��ĵ�����Ҫ�ɿ���������Bcl-2��Bcl-XL���µ��Լ��ٽ���������Bax��Bak��Bad���ϵ���ɡ����⣬ͨ���ϵ������ٵ�����������GOS2��DAP-3��DAP-3�����ŵ���øҲ���յ������IJ�����ά����D����ͨ������ͬԴ������ø(PTEN)�ı���������AKT���ڵĿ������ź�ͨ·�����ά����D��ͨ����ļCa2+�����ĵ�����������Ca2+�����Ħ�-�Ƶ���ø��Ca2+/�Ƶ���ø������ϸ����������ø-12�������������̡�
������һ����ϸ������͵���������ϸ����������������Ҫ���õķֽ��л���̡���Ȥ���ǣ��������ݱ���ά����D�ܹ�����ϸ���е�����ģʽ��ϸ������ת��������ά�����ܱ���ϸ�����Ի�������ϸ�����������ɣ������еķ����Ʒ�ͨ���յ�ϸ�����������ɡ�ά����D�յ���ϸ�������������̹�����һ�ֵ������ɵĻ���-beclin 1���ϵ������⣬CDK���Ƽ����ܰ�����ά����D���ڵ�ϸ���������������У���Ϊ������������ͨ��ʧȥp19�õ����ӻ�ʧȥp27����õ����������ң�ά����DҲ��ͨ���յ�DNA����-�յ�ת¼4(DDIT4)�͵��ڷ�����DNA���˵Ļ���(REDD1��һ���������ɵ�mTORC1���Ƽ�)�ı����������ɡ�
��֢��ά����D���Ե�ʧ��
ǰ���ᵽ�����͵Ĺǻ��������γ�øCYP27B1�������б�����ڹǻ��������ܺͽ����VDR��CYP24A1��һЩ�ض���֯���糦�������������б��Ȼ��������������֯����������λҲ�б��˵���ܶ�ϸ����������ά����D��Ŀ�겢�ҹǻ�������ˮƽ�ܹ��ֲ������Է��Ӿ�ϸ����֯���칦�ܡ���Ҫ���ǣ�����ά����D�ľֲ����������ںܶϸ���д���ʧ��״̬�������ڶԻ���ά����D�İ�֢���Ƶĵֿ������ڰ�֢�е�ά����D��л����ʧ�����ܽ�ɼ�ͼ4��
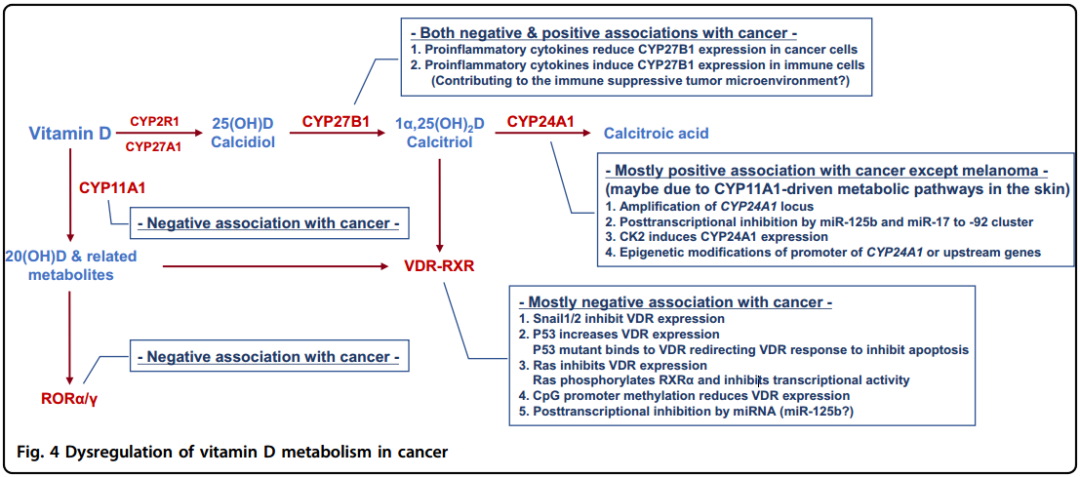
ͼ4��֢��ά����D�Ĵ�лʧ��
VDR/RXR��
VDR�ںܶ�ϸ���ж��б��VDR-RXRͨ���방֢��չ�ʸ���ع�ϵ���ں֢ܶ��VDR�ı��������ȥ�ֻ���������չ���͡�
��֢�Ľ�չһ�������ַ�ʽӰ��VDR�ı����һ��ת¼��������Snail�ں֢ܶ�ж����������Dz���EMT��������Ϯ��ת�ƣ���VDR�ı�����ֳ��������á��о�����Snail 1��Snail 2�ܹ���VDR���������������E-boxes�ϣ�Ȼ����ļ�谭�������ƽ�ֱ���������ٰ��е�VDRת¼���ڶ����������ƻ���p53���ڽ����������з�����ʧ��ͻ�䣬�ܹ�����VDR��ת¼����Ȥ���ǣ���֢��ص�p53ͻ��ϸ����ֱ����VDR����VDR����Ӧ������Ӳ���VDR���ڵ�ת¼���̷�ֹ��ϸ���������������֢ܶ�е��°�����Ras������ͻ�������VDR��������ֱ�����е�K-Rasͻ������С���ֱ������������Ƥϸ���е�H-Rasͻ�����������VDR��ת¼�Ӷ����ƹǻ�����������VDR�ļ�����˽���VDR�ı���ڽ����γ�ϸ����PECϸ��ϵ�зֱ���H-Ras��K-Ras��ͻ��������ͨ���յ�RXR�������ữ����VDR��ת¼�����ģ���֢��Ҳ�ᷢ��VDR�ı����Ŵ���Ĭ��VDR�����������CpG���������ֱ�������ٰ��е�VDR���������ء����ٰ�ϸ���е�DNA��ת��ø(DNMT)���Ƽ��յ�VDR�ı��ﲢ������˹ǻ������Ŀ���ֳЧ�������壬�б���������֢��miRNAҲ����VDR�ı��
CYP27B1
���ڹǻ�������ά����D����Ҫѭ����ʽ����ֻ������������ǻ�������ϸ����ȣ��ܱ���CYP27B1��ϸ�����һ�����ӹǻ������ľֲ�Ũ�ȡ���VDR���ƣ�CYP27B1�ı���Ҳ��ΰ���ǰ���ٰ�����ֱ��������״�ٰ���Ƥ�����Ľ�չ�ʸ���أ���������Щ��֯�йǻ������ľֲ�Ũ�ȿ��ܶ���ֹ��֢��չ����Ҫ���á���Ȥ���ǣ�Ŀǰ���о��������ڽ�ֱ�����У�����֢����IL-6��TNF-��ʹCYP27B1�ı����µ�����������֢��������Ҳ��������չ��ʹCYP27B1�����½���Ӱ�����ء����ǣ��������ַ��ӻ���Ŀǰ���������������Щ��֢�еĸ���������⣬�ڼ�״�ٰ������ٰ��������й۲쵽������صĽ������ΰ���ͬ���ǣ��ΰ����ߵķ��ݾ���ϸ���е�CYP27B1�����방֢��չ������ع�ϵ��������Բ��ֽ���Ϊ�ڵ���ϸ��������ϸ������ͻϸ���й۲쵽��������IFN-�á�TNF-����TLRsʹCYP27B1�����ϵ��������������������������Ӳ��鶯��ϸ����CYP27B1�ı����������ϸ����CYP27B1����ı仯��ά����D��л�Ĺ�ϵ��������������-��������������
CYP24A1
��֪CYP24A1��һ�ֿ��Խ���ǻ������ǻ�������ø����ô��ϸ������ͨ���ϵ�CYP24A1�ı��������ǻ������ľֲ�Ũ�ȣ����й�ϵ���ƺ֢ܶ��CYP27B1�Ľ��͡��ڴ�����о��У���ȷ����CYP24A1�ı����방֢��չ������أ����к�ɫ�����Ǹ����⡣
����CYP24A1���������ǻ�����-VDRͨ�������������յ��ģ���ϸ���и�ˮƽ��CYP24A1���ﲻ̫������VDR����鵼����Ϊ�ں֢ܶ�У�VDR�ı���ͻ��Զ��������µ�����˵���֢ܶ��CYP24A1�Ĺ����ﲻ�ǹǻ�����-VDR���Ƶĵ��ڽ����Ŀǰ�Ѿ���4�ֻ��Ʊ����������һ�����ٰ���һЩ������֢�е�CYP24A1�����������CYP24A1����Ⱦɫ��������йء���Ȥ���ǣ�ֻ��������֯�й۲쵽CYP24A1��������������֯����û�У�˵��CYP24A1���µĹǻ�����ʧ�����������״̬��չ�Ĺؼ����ء��ڶ�����֢�е�CYP24A1��������miRNAs��ת¼�������ء�������˿����/�հ��ᵰ��ø2(CK2)�ź�ͨ·����ǰ���ٰ��е�CYP24A1�������Ȥ���ǣ�CK2�ں֢ܶ�ж��ᷢ�����������ͨ���벻��Ԥ����ء����ģ������Ŵ�����CYP24A1����������Ҳ��ٽ���֢��CYP24A1����ĸı䡣
��Ƥ��ϸ���У�CYP11A1����ά����D�������л·����CYP24A1�ܲ�������ά����D��л�IJ�������ǽ���ǻ���������Ȥ���ǣ�CYP11A1������20(OH)D���Ա�CYP24A1�ǻ����γ�20,24(OH)2D��20,25(OH)2D�������ֶ��⻯����ǻ�������20(OH)D�Ժ�ɫ�����и��õ�����Ч���������һ�µ��ǣ�20-�ǻ�ά����D��л������ROR��/�õı������ɫ�����Ľ�չ�ʸ���ز��������õ�Ԥ����Щ�о�����������ά����D�������л·���������������Ƶ�һ���²��ԡ�
����ά����D�İ�֢����
��Ȼȱ��������ٴ�ʵ�����ݣ��ܶ����в�ѧ���ٴ����ٴ�ǰ������ʵ�����ݱ�������ά����D�ź��ǰ�֢Ԥ�������Ƶ���Ч���ԡ����ң��ܶ����ά����D��л�ļ�������ʧ���ĸ�Ԥ�����Ѿ����о����ڰ�֢���ơ�Ȼ�������Ʋ�������ЩDZ�ڵ�ȱ�ݻ��д����ơ�
ϵͳ�Լ���ά����D�źŵ�һ��DZ��Σ�����кܸߵĸ���������Ѫ��֢�����ܵ��º����ص��������ˡ�Ϊ�˽������ָ����ã��о���Ŭ����������VDR�Ŀ��������ǻ����������ά�ֿ������Ե�ͬʱ��С�����˸�Ѫ��֢�ļ��ʡ�ֱ�����ڣ��Ѿ�������Լ��1500��ά����D�����ֻ�к���һ���ֱ�֤���������ں����İ�Ѫ�������ٰ���ǰ���ٰ��ͽ᳦�����˵��ٴ������С���ΪCYP11A1������ά����D���ͨ·����Ҳ��Խ��Խ����˽��ˣ����Ƕ����ڰ�֢��VDR��ROR�еĹ��ܻ�֪֮���١�����·�����о��������ڿ�����֢�����е��²��ԡ����⣬��ϸ���йǻ������ľֲ�Ũ�ȵ���������һ�ֿ��Ա��������Ѫ��֢�IJ��ԡ���Ϊһ�ֹǻ���������ø��CYP24A1�ں֢ܶ�б���������������ø�������Ӱ�ϸ���йǻ������ľֲ�Ũ�ȡ���ȷ��Ŀǰ���о�����ͨ�������ó�����ҩ��ķ�ʽ����CYP24A1�Ļ��Կ��Ժܴ�̶����ӹǻ������Ŀ���Ч��������Ϊֹ�������ٴ�Ŀ��һЩCYP24A1�İ������Ƽ��Ѿ���������������Щ���Ƽ���֢�����Ƿ����ٴ���Ч���㹻�İ�ȫ�Ի��д��ٴ���֤��
���⣬���ڰ�֢��չ������CYP27B1��VDR��CYP11A1��ROR��/�õı�������ͣ�����ά����D������Ч�����ܾ��������ڽΡ���ˣ����ԺͿ����µ�����Ԥ�����ά����D��֢����Ч������ϱ������������ֲ��Ե��Գɹ�Ӧ�õIJ��ɻ�ȱ�IJ��֡����ң������о���֢��չ������Щø�ı��オ�͵Ļ���Ҳ�����еĹؼ����衣����ǰ���ᵽ��������ε��ز���CYP27B1-VDR��CYP11A1-ROR��/�ô�л���̵�ø�ı���Ļ������ǻ��˽��̫�١�Ŀǰ�������Ǻܶ�����ź�ͨ·ͨ��������Ӧø�ı�����л����Ը����˶�����лͨ·����Ӧ�ģ���Щ�������ź�ͨ·�ܴ�̶��ϵ���ά����D��л������ص�ø�ı���ͻ��ԡ��ҿ����ִ��۸��ӵİ������ά����D��лͨ·֮��Ĺ�ϵ��Ҫ��֢��ά����D��л���ܵ�ʧ���и���������⣬Ҳ�ܶ�֢�����ṩ����ϣ���IJ��ԡ�







