干货分享丨脂质组学入门篇
脂质定义
粗略的定义脂质是一类具有疏水性并且在大多数情况下可以溶于有机溶剂的物质,当然,还有部分脂质因为带有极性基团,往往是亲水性的,例如磷脂等。他们的生成、转运和识别是各种酶、结合蛋白和受体协同作用的结果。细胞的总脂质组成被称为脂质组,预计细胞中的脂质数目可能高达上万甚至上百万种,含量约在amol/mg到nmol/mg蛋白质水平。
脂质的分类方法多样,根据水解产物的复杂程度,可以将小于等于两个水解产物的分为简单脂质,大于三个或更多的分为复杂脂质;根据疏水性可以分为极性脂质和非极性脂质;根据功能可以分为膜脂质、能量脂质和生物活性脂质。为了促进脂质的国际交流,2005年,脂质代谢途径研究计划联合会(LIPID Metabolites And Pathways Strategy- LIPID MAPS)建立了一个对脂质进行全面分类的平台以便对海量的脂质数据进行信息学处理。
他们将脂质分为脂肪酰、甘油脂、甘油磷脂、鞘脂、固醇脂、异戊烯醇脂、糖脂和聚酮化合物八大类(图一)。根据脂质内部不同结构单元的组合和功能可以对脂质进行细分,目的在于拓展脂质的种类,在这个分类中,几何位置的标记可以区分不同的亚类并且对某些特殊官能团进行定位,例如,在甘油脂中,sn-1(立体定向编号)的氧原子通过酯键、醚键或烯醚键等方式与取代基进行结合;“d”和“t”分别指代鞘脂中1,3-二羟基和1,3,4-三羟基长链的碱基;双键的几何构型通常用E/Z表示;“溶血(lyso)”表示在甘油类的脂中缺少一个酰基、O-烷基或O-烯基等基团。
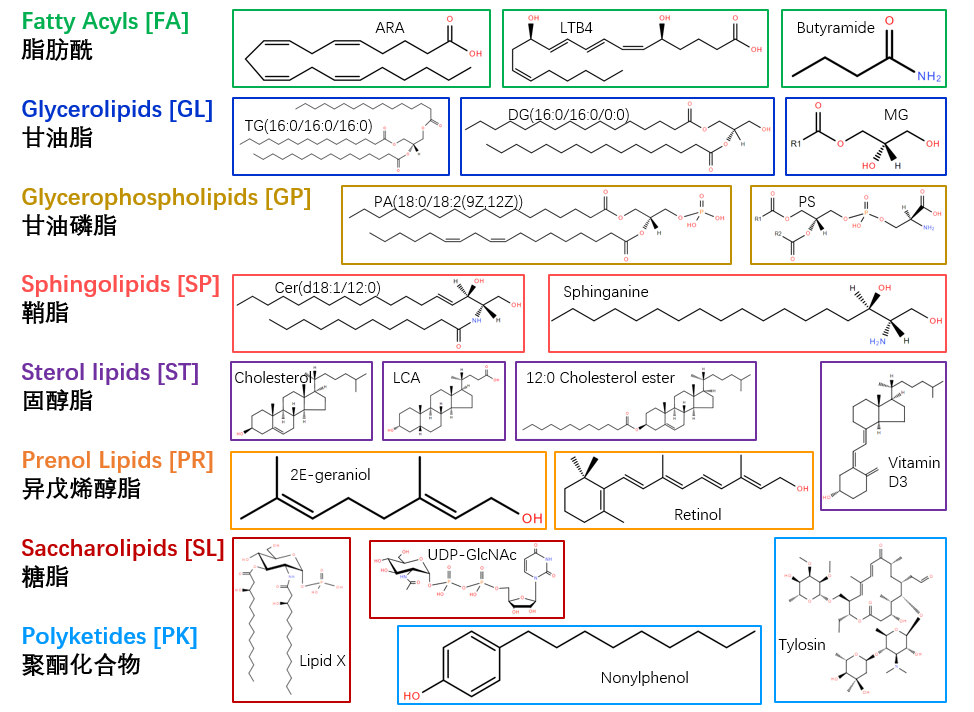
图一 八类脂质结构示例
脂肪酰(FA)是以乙酰辅酶A作为引物,与可能含有环状结构或者杂原子的乙酰基辅酶A或丙二酰辅酶A通过链长延伸的方式合成的一类分子,包括游离脂肪酸和类花生酸等。
含有甘油基团的脂质有两类:甘油脂(GL)和甘油磷脂(GP),它们的区别在于与甘油上的羟基进行酯化的是脂肪酸还是磷酸。甘油脂的核心结构甘油上的三个羟基可以分别被取代形成甘油单酯(MAG)、甘油二酯(DAG)和甘油三酯(TAG)。
甘油脂(GL)本质上包含所有含甘油的脂质,但由于甘油磷脂(GP)的高丰度、是膜结构的重要组成成分、参与能量代谢和信号传导而将其单独分类。此外,根据甘油磷脂的极性头基的不同取代形式可以对GP进行分类。例如,根据图二中X取代的不同可以分为胆碱磷脂(PC)、乙醇胺磷脂(PE)、甘油醇磷脂(PG)、肌醇磷脂(PI)等。而甘油的sn-1位的羟基被取代形成磷脂酰基-、磷脂醚-或缩醛磷脂-等亚类。

图二 甘油磷脂的分类
鞘脂(SP)是由丝氨酸和长链的脂肪酰辅酶A从头合成的一类复杂的化合物,它们的核心结构是长链鞘氨醇碱基。一些与鞘脂有部分相似特征并且能够产生鞘氨醇碱基类似物的也被归为鞘脂,因为它们在某些生物体中可以作为鞘脂的抑制剂或抗结剂而在另一些生物体中可以作为鞘脂的替代物。鞘脂的骨架结构鞘氨醇也有三个取代位置,其中C1位的氧可以连接不同的极性基团,包括磷酸乙醇胺、鞘磷脂、半乳糖等,C2位的氨基可以和各种脂发生结合形成脂肪酰基,C3位的碱基可以连接不同长度和饱和度的烷基链(图三)。

图三 鞘脂的核心结构及代表类型
固醇脂(ST)的核心结构是四个稠环(图四),胆固醇及其衍生物、类固醇、胆汁酸是哺乳动物中被研究的较多的类型。根据核心结构上的碳原子数可以进一步分类,C18固醇包括雌激素类,C19固醇包括雄性激素,C21固醇包括孕激素等。

图四 固醇脂的核心结构
异戊烯醇脂(PR)是由异戊烯二磷酸酯和二甲基烯丙基二磷酸酯通过甲羟戊酸等路径合成,包含类异戊二烯、醌类等化合物。
糖脂(SL)是一类脂肪酰基直接和糖骨架相连的脂质,糖取代了甘油脂和甘油磷脂中的甘油骨架。最常见的糖脂是革兰氏阴性菌中脂质A的乙酰化葡萄糖胺前体,最小的脂多糖是用于E. coli生长的已糖酰化的脂质A。在某些细菌中,脂质A的骨架可以被含氨基的化合物取代形成“酰化糖胺”。
最后一类脂质是聚酮化合物(PK),它的合成需要酶和酰基载体蛋白的参与,而且聚酮合成酶会产生更多的具有脂质特性的天然产物。聚酮的骨架结构可以进一步被糖基化、羟基化、甲基化等各种修饰。
此外,在这个分类方法中,每个脂质分子在LIPID MAPS中都有唯一的12位编码,这个编码包含了数据库来源、每个分子的类、亚类,系统名称、分子式等信息(图五)。

图五LIPID MAPS中脂质分子的信息
脂质功能
脂质能够直接反映细胞的代谢状态,它们不仅是细胞膜的重要组成部分,而且具有能量存储、信号传导、促炎症或抗氧化等多种不同的生物学功能。大多数极性脂质是生物体内生物膜的重要组成成分,比如PC、PE、PS、PI、PA、SM与Cho是真核细胞膜上的主要脂质结构。同时,脂质也是生物体内能量存储与释放过程中的关键物质。当能量过多且需要储存时,三羧酸循环产物乙酰辅酶A可通过从头合成途径产生不同链长脂肪酸,并通过甘油骨架、鞘氨醇骨架或游离胆固醇结合,最终生成TAG、PC、与CE等各种脂质亚类。因此,能量代谢相关的脂质含量变化已是肥胖、糖尿病、脑卒中等多种疾病研究的重要关注方向。许多脂质也可作为重要第二信使或前体物质来参与生物体内信号传导。FAs、GPs、GLs与SPs通过各种水解酶释放脂肪酸的同时,还会水解产生LPC、LPA、IP3、DAG、SPH与S1P。除此之外,缩醛磷脂作为内源性抗氧化剂,能够有效抵抗PUFA通过自由基的非酶促氧化或脂氧合酶的酶促氧化引起的细胞铁死亡。一些聚酮或聚酮衍生物可以作为杀菌剂或者抗癌药物,还有一些是潜在的毒素。因此,脂质组成与生理或病理过程变化密切相关。

图六 脂质功能
脂质鉴定
很多工具例如GC、HPLC、TLC、MS、NMR等都曾用于脂质的结构鉴定、发现和定量。而脂质分析的突破性进展获益于1980s年代软电离源(ESI和MALDI)的发展,这两种离子源的发展实现了对完整脂质分子的定性定量。1990s年代,关于这两种离子源在脂质研究的应用开始陆续发表。典型的例如Henion等人用直接灌注的方式分析单甘油脂、双甘油脂和甘油三脂的混合标准品,他们发现在软电离源中脂质的源内裂解碎片最少,分子离子的响应与脂类的极性和双键数目相关,离子化效率随着酰基链的长度增加而降低,而且分子离子中经常会产生含钠或者含胺的加合物。随着色谱和质谱串联技术的进一步发展,加速了对低丰度脂质的定性定量和对复杂脂质的深度鉴定。关于这一部分的内容将后续文章中进行详细介绍。
参考文献
[1] Han X, Gross R W. The foundations and development of lipidomics[J].Journal of lipid research, 2022: 100164.
[2] Fahy E, Subramaniam S, Brown H A, et al. A comprehensive classification system for lipids1[J].Journal of lipid research, 2005, 46(5): 839-861.
[3] Fahy E, Subramaniam S, Murphy R C, et al. Update of the LIPID MAPS comprehensive classification system for lipids1[J].Journal of lipid research, 2009, 50: S9-S14.
[4] Züllig T, Köfeler H C. High resolution mass spectrometry in lipidomics[J].Mass Spectrometry Reviews, 2021, 40(3): 162-176.
[5] Han X. Lipidomics for studying metabolism[J].Nature Reviews Endocrinology, 2016, 12(11): 668-679.
[6] Schramp M, Hedman A, Li W, et al. PIP kinases from the cell membrane to the nucleus[J].Phosphoinositides I: Enzymes of Synthesis and Degradation, 2012: 25-59.
[7] Cui W, Liu D, Gu W, et al. Peroxisome-driven ether-linked phospholipids biosynthesis is essential for ferroptosis[J].Cell Death & Differentiation, 2021, 28(8): 2536-2551.







