经典重读丨Cell Metab细胞脂肪酸代谢与癌症
癌细胞通常具有代谢的特征性变化。细胞增殖是所有癌症的共同特征,细胞增殖需要脂肪酸来合成膜和信号分子。加利福尼亚大学心血管疾病研究所的Erin Currie等人在2013年6月在Cell Metabolism期刊上发表题为“Cellular Fatty Acid Metabolism and Cancer”的文章,从脂质的角度对癌细胞代谢进行了综述,并总结了限制脂肪酸可用性来控制癌细胞增殖的依据。原文链接:http://dx.doi.org/10.1016/j.cmet.2013.05.017。

前言
尽管癌症在类型和病因上有很大的差异,但癌细胞通常具有代谢异常的特征。例如,葡萄糖代谢通常会发生改变,即使在氧气充足的条件下,癌细胞糖酵解一样活跃(Warburg效应)。而癌细胞中脂肪酸(FA)代谢的变化较少受到关注,FAs由末端羧基和烃链组成,主要以偶数碳的形式存在,可以是饱和的,也可以是不饱和的。它们是能量储存、膜增殖和信号分子合成所必需的。在这里,我们简要回顾癌细胞中的代谢,重点是FA合成和储存的途径。此外,我们还研究了一种通过限制FA可用性来减弱癌细胞增殖和转移的模型。
癌细胞能量代谢的改变
癌症从根本上来说是一种细胞生长和增殖的紊乱行为,它需要细胞的基本构件,如核酸、蛋白质和脂质。癌细胞的新陈代谢常常受到干扰,这使得它们能够积累新陈代谢中间产物,作为这些构件的来源。癌细胞中最易理解的代谢紊乱是Warburg效应,即使在有氧的情况下也使用糖酵解途径,其特征是葡萄糖摄取和消耗增加,氧化磷酸化减少,以及乳酸的产生。
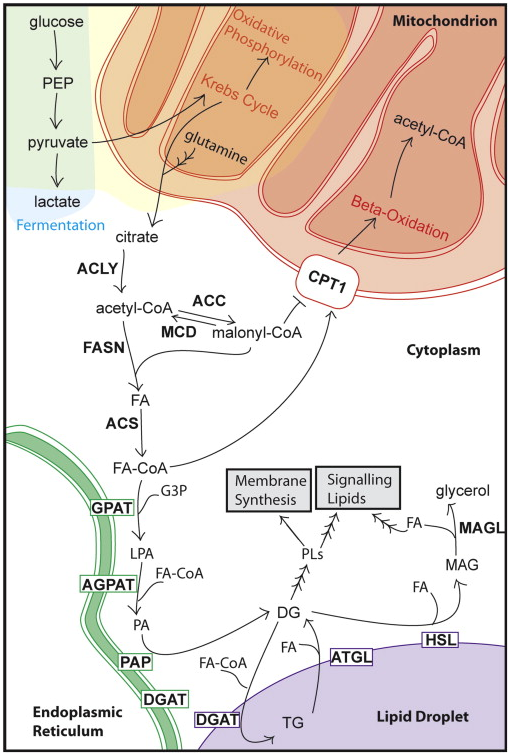
图1细胞脂肪酸代谢概述
另一个常见的癌症代谢改变是谷氨酰胺代谢增加。在哺乳动物细胞中,谷氨酰胺是一种主要的能量底物,经过代谢可以转化为α-酮戊二酸,从而进入三羧酸循环。谷氨酰胺衍生的α-酮戊二酸通过三羧酸循环的正向运转会产生苹果酸酶依赖的丙酮酸,有助于柠檬酸的产生。谷氨酰胺也可以在异柠檬酸脱氢酶和乌头酸酶催化作用下,经三羧酸循环的逆向运转产成柠檬酸,然后柠檬酸可用于生产脂肪酸合成的乙酰基。
快速增殖细胞中的脂质代谢也发生改变。在癌细胞中,碳必须从能量生产转移到脂肪酸,用于膜和信号分子的生物合成。大部分细胞膜脂质是磷脂(PLs),如磷脂酰胆碱(PC)和磷脂酰乙醇胺(PE),以及其他脂质,如甾醇、鞘磷脂和溶酶体磷脂。这些脂质许多含有脂肪酸,部分脂质来源于乙酰辅酶A。脂肪酸构件来自外源或从头合成的脂肪酸。虽然大多数正常细胞更喜欢外源性来源,但癌细胞偏好从头合成脂肪酸,并经常表现出向脂肪酸合成的转变。为了进入生物活性池,脂肪酸需要通过脂肪酰基辅酶A合成酶的共价修饰进行“激活”。一旦进入活性池,脂肪酸可以与甘油或甾醇骨架酯化,分别生成三酰甘油(TGs)或甾醇酯(SEs),然后储存在脂滴(LDs)中(见图1)。在细胞内,脂肪酸可能有许多命运,包括被结合到膜上、被储存、变成信号脂质、被氧化成二氧化碳作为能量来源。
尽管本综述侧重于从头开始的脂肪酸合成途径,但一些癌症从其环境中清除脂质,使脂肪酸摄取途径成为潜在的靶点。例如,脂肪酸结合蛋白4(FABP4),是一种脂质伴侣,参与从周围脂肪细胞向卵巢癌细胞提供脂肪酸的过程。此外,只有在不存在脂蛋白(一种外源性脂质源)的情况下培养时,前列腺癌细胞在存在FASN(C75)或ACLY(SB-204990)抑制剂的情况下显示出生存能力降低。CD36是一种广泛表达的跨膜蛋白,具有多种功能,包括脂肪酸摄取,与乳腺癌有关,基质组织中CD36水平降低与肿瘤发生的早期步骤相关。值得注意的是,细胞培养实验的体外条件可能不同于体内条件,其中外源性摄取在某些癌症中可能更为重要。
通过限制脂肪酸供应来限制癌细胞增殖
由于脂肪酸对癌细胞增殖至关重要,限制其可用性能提供一种潜在治疗策略。从脂质代谢的角度来看限制脂肪酸可用性可以通过几种方式实现:(1)阻断脂肪酸合成;(2)增加脂肪酸氧化分解;(3)将脂肪酸转移到存储池中;(4)阻止存储池中的脂肪酸的释放(图2)。这些限制方式可以单独或以组合方式完成,以此为框架,我们回顾了与该模型相关的证据。
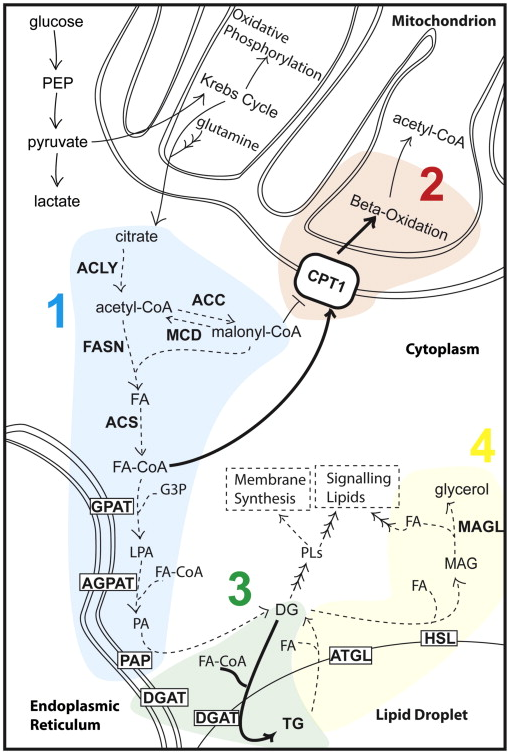
图2通过限制脂肪酸来限制癌细胞增殖的模型
阻断脂肪酸合成
降低脂肪酸水平的最简单方法是阻断它们的合成。葡萄糖代谢成丙酮酸后,通过三羧酸循环生成柠檬酸,从而进入脂肪酸代谢(见图1)。碳从柠檬酸转化为生物活性脂肪酸需要几个步骤,这些步骤涉及ATP柠檬酸裂解酶(ACLY、ACL或ATPCL)、乙酰辅酶A羧化酶(ACC)、脂肪酸合成酶(FASN或FAS)和酰基辅酶A合成酶,也称为脂肪酸辅酶A连接酶(ACS、ACSL或FACL)。在降低脂肪酸可用性的模型中,抑制这些酶能限制癌细胞生长。这些策略的临床意义很重要,因为这些酶的抑制剂对非癌细胞的影响最小。
柠檬酸的亚细胞定位决定了它的代谢命运:线粒体中的柠檬酸会进入三羧酸循环,细胞基质中的柠檬酸会加入脂肪酸合成。转运蛋白CIC(柠檬酸盐载体)以调节的方式转运柠檬酸穿过线粒体内膜以在细胞基质中使用。CIC水平在各种癌症中与不良预后正相关,苯三羧酸类似物(BTA)对转运的抑制在各种肿瘤类型和异种移植小鼠体内显示出抗癌作用。
阻断脂肪酸合成基因的表达
除了直接靶向抑制脂肪酸合成酶外,也可以通过降低转录水平来降低脂肪酸水平。脂肪酸合成的主要转录调控因子是甾醇调节元件结合蛋白1(SREBP-1)。SREBP-1有两种亚型:SREBP-1a是大多数培养细胞系中的主要亚型,SREBP-1c在肝脏和大多数组织中占主导地位。在正常水平下,SREBP-1c通过应答基因(包括ACLY、ACC、FAS、SCD-1和GPAT)激活脂肪酸生物合成途径。因此,抑制癌细胞中的SREBP-1可以降低脂肪酸合成基因的表达,并能阻止癌细胞增殖。在上游,SREBP转录因子受多个信号通路调节,包括生长因子信号传导和肝脏X激活受体(LXR),通过诱导SREBP-1c表达来激活脂肪酸合成。因此,可以通过抑制LXR激活来减弱癌细胞增殖。然而,LXR的激活,特别是通过T0901317,反而抑制了乳腺癌、结肠癌和前列腺癌中癌细胞的增殖,这些说明LXR除了调节FA合成之外还有功能。
增加脂肪酸氧化分解
降低癌细胞中脂肪酸水平的第二个方法是增加其降解速率。激活的脂肪酸能被线粒体β氧化所代谢。脂肪酸辅酶A通过肉碱棕榈酰转移酶1(CPT1)转化为脂肪酸肉碱后,从胞浆跨线粒体外膜转运。在线粒体内,每进行一次β-氧化作用,会分解出一个二碳片段,生成较原来少两个碳原子的脂肪酸脂,产生乙酰辅酶A,然后进入三羧酸循环释放大量ATP。增加脂肪酸氧化以限制脂肪酸水平在理论上可行,但实验数据很难验证。
将脂肪酸转移到存储池中
脂肪酸可用于膜脂合成、降解或储存。可以想象,中性脂质(如TGs或甾醇酯)中脂肪酸的储存增加后,可用作膜构建或信号脂质的活性脂肪酸会减少,从而抑制细胞增殖。大多数细胞将脂肪酸储存在甘油三酯中的胞质脂滴(LD)中,该细胞器的主要功能是脂质储存。LDs在癌细胞中的作用尚不清楚。虽然据报道,许多癌细胞中胞质脂滴数量增加,并且这种累积被认为是致病性的,但胞质脂滴的累积本身可能不是罪魁祸首。它们所代表的现成的脂肪酸池可能是致病性的。胞质脂滴累积也可能反映细胞对应激的反应。未来的研究还应仔细描述胞质脂滴累积是发生在癌细胞内还是周围细胞内。
主要的甘油三酯合成途径称为Kennedy或磷酸甘油途径。它使用甘油-3-磷酸酰基转移酶(GPAT)、酰基甘油磷酸酰基转移酶(AGPAT)、磷脂酸磷酸水解酶(脂蛋白或PAP)和二酰基甘油酰基转铁酶(DGAT)将脂肪酸与甘油3-磷酸缩合。除最远端酶(DGAT)外,所有酶的产物都进入磷脂合成。因此,GPAT、AGPAT和脂蛋白可能被抑制以限制磷脂产生,而增加脂肪酸储存的努力将集中于激活DGAT。此外,增加脂肪酸储存的潜在益处可能只有在同时抑制脂肪酸从储存中释放的情况下才能实现。
阻止存储池中的脂肪酸的释放
一旦储存,脂肪酸可被释放供特定脂肪酶使用。通过阻止存储池中脂肪分解,可用于癌细胞增殖的活性脂肪酸池可能减少。脂肪分解产生的脂肪酸也可作为重要信号脂质的前体。大多数关于脂肪分解的知识来源于对脂肪细胞的研究,其中乳酸脱氢酶(LD)中的每个甘油三酯分子可通过脂肪甘油三酯脂肪酶(ATGL)、激素敏感脂肪酶(HSL)和单酰基甘油脂肪酶(MAGL)的顺序作用被完全水解释放三种脂肪酸。尽管这些脂肪酶中的每一种在其他组织中也具有重要功能,但尚不清楚其他脂肪酶是否在其他细胞类型中发挥作用。目前,大多数关于脂肪酶和癌症的数据都是针对单酰基甘油脂肪酶的。
结论
癌细胞依赖脂肪酸作为细胞基本构件,用于膜形成、能量储存和信号分子的产生。如果癌细胞增殖需要脂肪酸的模型是正确的,那么癌细胞可能在脂肪酸代谢途径中存在多个靶点,可以破坏癌细胞快速增殖,而这些靶点已经存对应的抑制剂(表1)。就像葡萄糖代谢一样,针对脂肪酸代谢可能对高增殖细胞更具选择性。或者,以细胞特异性和靶向性的方式来递送脂肪酸代谢抑制剂也是潜在治疗方式。
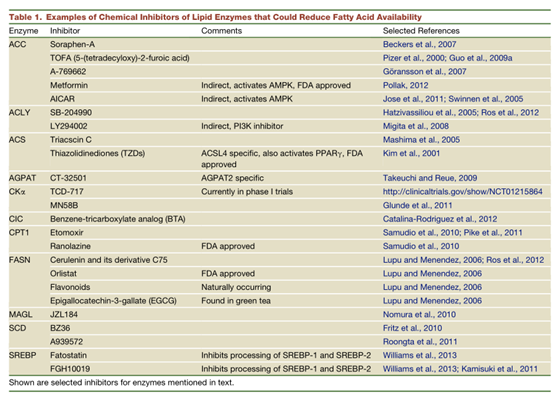
癌症的类型和潜在的基因改变是多种多样的。脂质代谢更是复杂的,有许多不同的反馈机制和调节点。此外,大多数脂质代谢酶具有多种亚型,这些亚型可与不同的脂质代谢过程耦合,具有不同的细胞定位或组织分布。因此,治疗方式的成功与否可能取决于对特定类型癌症的特定代谢异常机制的了解程度。







